Lack of Juvenile Photorefractoriness in Japanese Quail
by Emily Stachowski1, Andrew Benson2
1 UGA Warnell School of Forestry & Natural Resources
2 UGA College of Agricultural & Environmental Sciences,

Photoperiod is a critical environmental cue regulating reproductive development in birds through the activation of the hypothalamic-pituitary-gonadal (HPG) axis. This study investigated the effects of varying durations of photostimulation on Japanese quail (Coturnix japonica) body weight, testicular development in males, and ovarian and oviductal development in females. A total of 150 quail, divided equally by sex, were assigned to three photoperiod treatments: 0-weeks (non-stimulatory light for 7 weeks), 2-weeks (non-stimulatory light for 5 weeks followed by 2 weeks of stimulatory light), and 7-weeks (stimulatory light for the full duration). Body weights, testicular weights in males, and ovarian and oviductal weights in females were measured at the end of the seven weeks. The results indicated no significant difference in body weight among males across treatment groups, whereas females in the 7-week group had significantly higher body weights compared to the other groups. Testicular weights in males and ovarian and oviductal weights in females were significantly greater in the 7-week group compared to the 0 and 2-week groups. These findings suggest that Japanese quail do not experience juvenile photorefractoriness and provides insights into the role of photoperiod in optimizing reproductive maturation, with potential applications in avian breeding programs.
Japanese quail, juvenile photorefractoriness, photostimulation, photoperiod, gonadal growth
Introduction
Juvenile photorefractoriness in birds refers to the temporary physiological state during which young birds are unresponsive to increasing day length (photoperiod) that would normally stimulate reproductive development1. In this refractory state, juvenile birds remain unaffected by increased exposure to daylight, which typically activates the hypothalamic-pituitary-gonadal (HPG) axis, leading to gonadal growth and maturation.
Photorefractoriness is part of an evolutionary adaptation that ensures that birds do not attempt to reproduce during unfavorable seasons, such as winter, when conditions for raising offspring would be poor2. In species like European Starling, juvenile photorefractoriness prevents early reproductive maturation until the birds have reached an age and developmental stage where they can support reproductive processes1. Once this refractory period passes, birds become responsive to increasing photoperiods and will begin to undergo gonadal development as they prepare for the breeding season3.
Juvenile photorefractoriness ensures that young birds prioritize growth and survival before entering the energetically demanding reproductive phase of their life cycle. In controlled environments, such as poultry production systems, the timing of photostimulation is carefully managed to break this refractory state and induce reproductive activity at the desired age4.
The Japanese quail (Coturnix japonica) is widely regarded as an important model species for studying the effects of photoperiod on avian reproduction due to its well-characterized reproductive physiology, rapid development, and ease of maintenance in laboratory settings. This species has a short generation time and reaches sexual maturity within 5-6 weeks, allowing for the efficient study of reproductive cycles within a relatively short timeframe. Japanese quail are highly responsive to changes in photoperiod, which makes them particularly useful for investigating the mechanisms by which light influences the HPG axis and regulates reproductive development. The quail’s sensitivity to photoperiod also makes them an excellent model for exploring juvenile photorefractoriness, seasonal breeding behaviors, and the interaction between light and endocrine signals that govern reproductive cycles. As such, findings from studies using Japanese quail often have broader implications for understanding photoperiodic regulation in other bird species, including those of ecological and agricultural importance, and contribute to insights into avian reproductive physiology. Our experiment’s objective was to evaluate photostimulation’s effects on juvenile photorefractoriness in Japanese quail and determine if they experience this phenomenon.
Methods
Japanese quail were grown in battery cages for seven weeks starting on day one after hatching at the University of Georgia Poultry Research Center. The Japanese quail were maintained under appropriate brooding conditions with ad libitum access to food and water in 18-inch-high battery cages at a stocking density that allowed 0.5 square feet per bird. Quail were assigned to one of three treatment groups consisting of 50 birds each. In the 0-week photostimulation group, birds were housed under non-stimulatory light conditions, consisting of six hours of light and 18 hours of dark for the full seven weeks. In the 2-week photostimulation group, birds were housed under non-stimulatory light conditions for five weeks (6 hours of light and 18 hours of dark) and then exposed to stimulatory light conditions (18 hours of light and 6 hours of dark) for the final two weeks. In the 7-week photostimulation group, birds were housed under stimulatory light conditions (18 hours of light and 6 hours of dark) for the entire seven weeks. At the end of the experiment, birds were euthanized using CO2 gas, and body weights were recorded on Veritas M124A electronic scales.
Following euthanasia, necropsies were performed to collect reproductive organs. Testes were removed from males and weighed on electronic scales, while ovaries and oviducts were removed from females and weighed. One-way ANOVA was used to analyze body weight, ovarian weight, oviduct weight, and testicular weight across the three treatment groups, followed by post-hoc analyses with Tukey’s HSD to determine pairwise differences, with statistical significance set at p < 0.05. All animal procedures were approved by the Animal Care and Use Committee of the University of Georgia (A2022 04-004-Y3-A1).
Results
There was no significant difference (p=0.34) in body weight among the treatments in males with 0-weeks averaging 205.6g, 2-weeks averaging 216.2g, and 7-weeks averaging 208.2g (Figure 1). There was, however, a significant increase in body weight in females in the 7-weeks photostimulation treatment (p=0.01) averaging 245.9g but not among 0-weeks averaging 218.4g and 2-weeks averaging 225.1g (Figure 2). The oviduct and ovary weights also significantly increased in females among the 7-weeks of photostimulation treatment (p=0.01 & p<0.01) averaging 4.1g oviducts and 3.4g ovaries; however, there was not a difference between 0-weeks averaging 0.05g oviducts and 0.09g ovaries and 2-weeks averaging 0.15g oviducts and 0.23g ovaries (Figures 3-4). In the males, the testes’ weight increased significantly (p <0.01) for the 7-weeks of photostimulation treatment averaging 4.4g, and significantly increased between the treatment of 0-weeks of photostimulation averaging 0.06g and the treatment of 2-weeks averaging 1.3g (Figure 5).
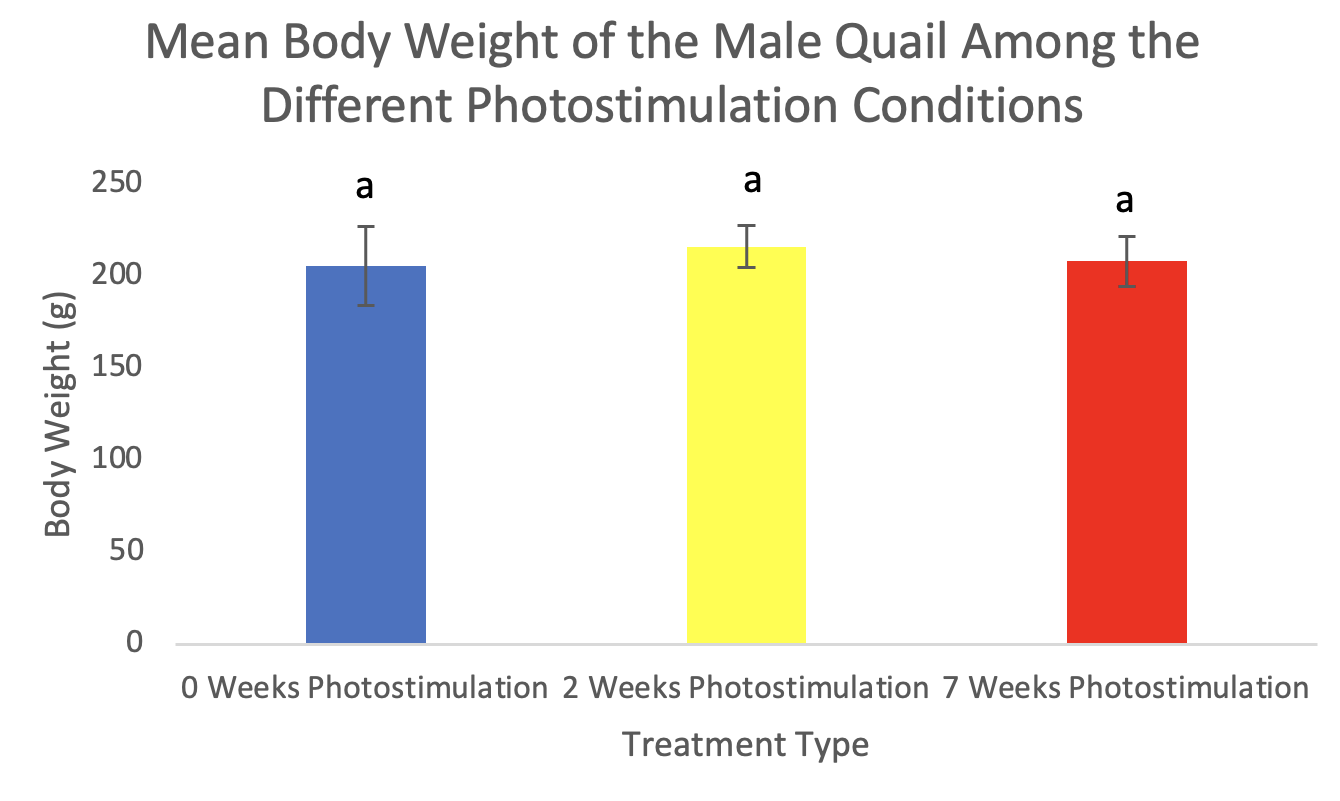
Figure 1. Body weights (mean ± standard deviation) of male Japanese quail. Values are means ± SEM. There was no significant difference between groups (p = 0.34).
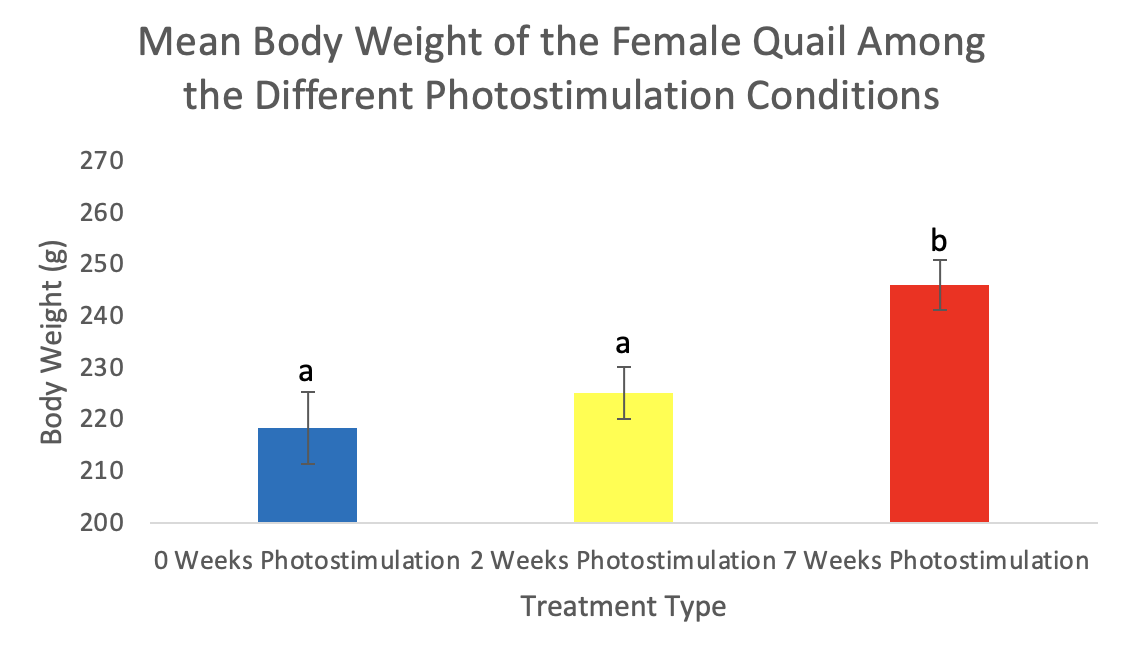
Figure 2. Body weights (mean ± standard deviation) of female Japanese quail. Values are means ± SEM with different letters indicating significant differences (p <0.01).
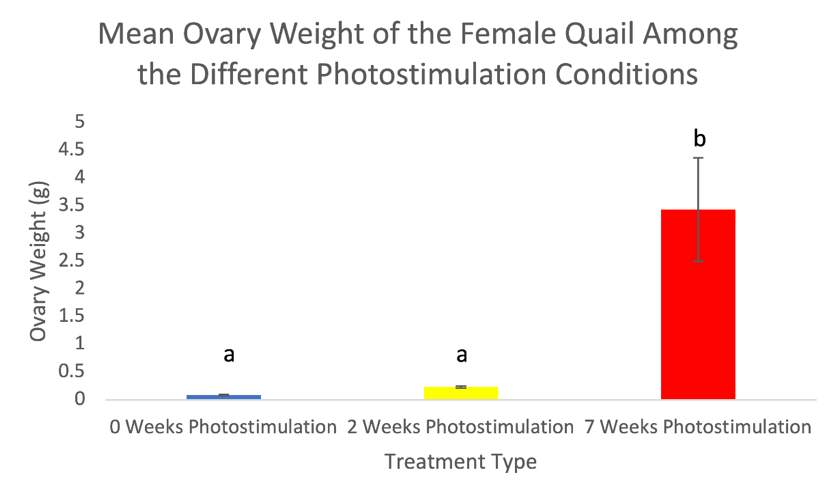
Figure 3. Ovary weights (mean ± standard deviation) of female Japanese quail. Values are means ± SEM with different letters indicating significant differences (p <0.01).
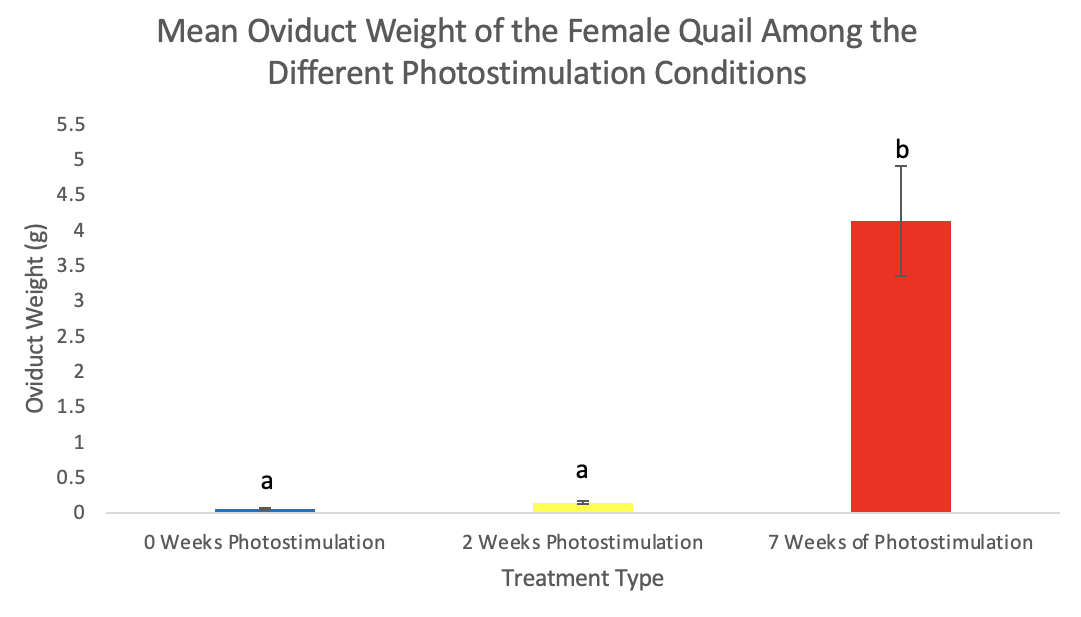
Figure 4. Oviduct weights (mean ± standard deviation) of female Japanese quail. Values are means ± SEM with different letters indicating significant differences (p <0.01).
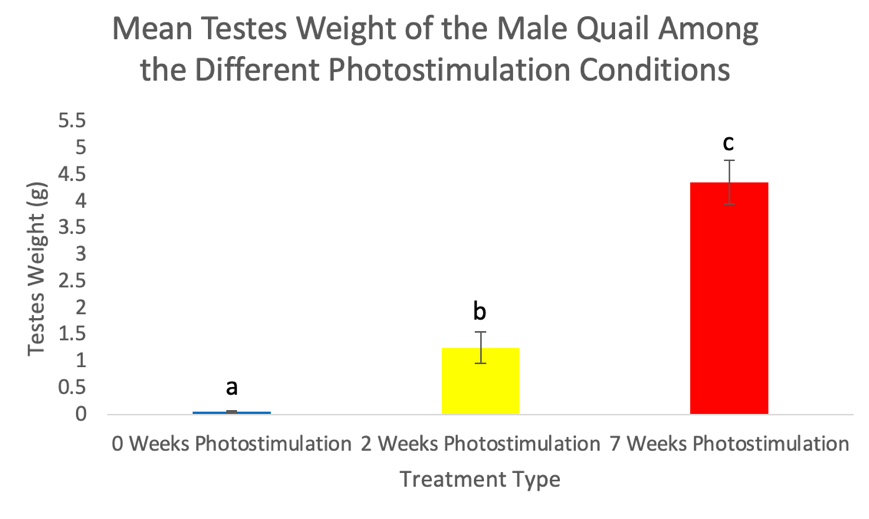
Figure 5. Testes weights (mean ± standard deviation) of male Japanese quail. Values are means ± SEM with different letters indicating significant differences (p <0.01).
Discussion
The results of this study demonstrate a clear influence of photostimulation on reproductive organ development in both male and female Japanese quail, while body weight was only significantly affected in females. In males, no significant difference was observed in overall body weight among the treatment groups. This suggests that photostimulation did not influence somatic growth in male quail over the seven weeks, which is consistent with previous studies showing that body mass and reproductive organ development can be independently regulated in birds5. However, the significant increase in testes weight in the 7-week photostimulation group compared to the 0 and 2-week groups indicates that prolonged exposure to stimulatory light conditions promotes gonadal development in males.
In females, the results showed that birds in the 7-week photostimulation group had significantly greater body weights than those in the 0 and 2-week groups. This difference could be attributed to the energy demands associated with ovarian and oviductal development6, which were also significantly greater in the 7-week group. The increase in ovarian and oviductal mass in response to extended photostimulation highlights the sensitivity of the female reproductive system to photoperiod changes, as longer daylight exposure triggers the development of these reproductive tissues in preparation for egg production.
The data suggest that seven weeks of continuous photostimulation can promote full reproductive maturation in both male and female Japanese quail, as evidenced by the significantly greater reproductive organ weights in this group. Shorter periods of photostimulation (two weeks) were not as effective, indicating that a minimum duration of light exposure is necessary to induce maximal reproductive organ development. These findings align with established knowledge of photoperiodic control of avian reproduction, wherein increasing day length activates the HPG axis, leading to gonadal growth and reproductive readiness4. Importantly, this study demonstrates that Japanese quail are not restrained by the need for a scotophase period; therefore, they do not possess the more widely held juvenile photorefractory state evident in most avian species7.
In conclusion, the study highlights the importance of photostimulation in optimizing reproductive output in Japanese quail. While body weight was unaffected in males, females exhibited increased body mass under prolonged light conditions, likely due to the energy demands of reproductive tissue growth. Further research could explore the hormonal mechanisms underlying these responses and investigate the long-term effects of varying photoperiods on reproductive success and overall health in quail.
Works Cited
1 McNaughton, F. J., et al. “Juvenile Photorefractoriness in Starlings, Sturnus vulgaris, Is Not Caused by Long Days After Hatching.” Proceedings of the Royal Society of London. Series B: Biological Sciences, vol. 248, no. 1322, 22 May 1992, pp. 123-128, https://doi.org/10.1098/rspb.1992.0051.
2 Hahn, T. P., et al. “Evolution of Environmental Cue Response Mechanisms: Adaptive Variation in Photorefractoriness.” General and Comparative Endocrinology, vol. 163, no. 1-2, 23 Apr. 2009, pp. 193-200, https://doi.org/10.1016/j.ygcen.2009.04.012.
3 Lewis, P. D., et al. “Photoperiods for Broiler Breeder Females during the Laying Period.” Poultry Science, vol. 89, no. 1, 1 Jan. 2010, pp. 108-114, https://doi.org/10.3382/ps.2009-00312.
4 Benson, A. P., et al. “Effect of Early Photostimulation at 15-Weeks of Age and Everyday Spin Feeding on Broiler Breeder Performance.” Poultry Science, vol. 101, no. 6, June 2022, 101872, https://doi.org/10.1016/j.psj.2022.101872.
5 Guioli, S., et al. “Gonadal Asymmetry and Sex Determination in Birds.” Sexual Development, vol. 8, no. 5, 27 Feb. 2014, pp. 227-242, https://doi.org/10.1159/000358406.
6 Williams, T.D., et al. “The Hormonal and Physiological Control of Egg Production.” OUP Academic, Oxford UP, 5 Aug. 2012, academic.oup.com/princeton-scholarship-online/book/23234/chapter-abstract/184149128?redirectedFrom=fulltext.
7 Watts, S., et al. Doing Q Methodological Research: Theory, Method and Interpretation. SAGE, 2012.
Acknowledgements: We thank Jesse Hanks for rearing the quail. We thank Dr. Joe Bartges, Jacob Knox, and the Classic Journal team for helping in the editing and publication process.
Citation Style: MLA